La tòfona és el cos de fructificació d’un fong hipogeu ectomicorízic, és a dir, que produeix les seves espores en carpòfors sota terra i que s’associa a una planta hoste de la qual rep carbohidrats.
La planta hoste també es beneficia de la simbiosi, ja que rep aigua i minerals del fong, especialment quan creix en sòls pobres en fòsfor. En aquesta simbiosi, el fong creix al voltant i entre les cèl·lules externes de les arrels tròfiques formant una estructura coneguda com a micoriza que permet l’intercanvi de substàncies entre ambdues espècies. Els principals hostes que s’utilitzen actualment per al seu cultiu són les alzines (Quercus ilex ssp. ilex, Q. ilex ssp. ballota), els roures (Q. faginea, Q. pubescens), els garrics (Q. coccifera) o els avellaners (Corylus avellana).


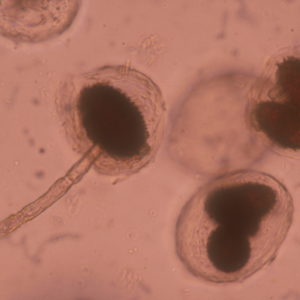
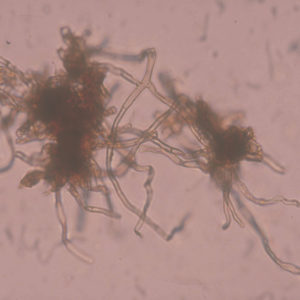
La tòfona negra es caracteritza per formar en el terreny una zona desproveïda de vegetació al voltant de l’arbre, anomenada cremat . Els cremats són una mostra de la capacitat fitotòxica d’aquest fong per crear un ambient que li sigui més favorable, com per exemple, reduir la competència per l’aigua del sòl per part la vegetació herbàcia o afavorir l’acumulació del carbonat càlcic.
Hi ha poques plantes que resisteixen el cremat. Entre elles, el cirerer de Santa Llúcia (Prunus mahaleb), el sanguinyol (Cornus sanguinea), els càdecs i ginebres (Juniperus oxycedrus, J. communis), el crespinell gros (Sedum altissimum) o el pèl de guilla (Festuca rubra), entre d’altres.
Durant els últims 10 anys s’han publicat molts estudis científics dedicats al coneixement del cicle biològic de la tòfona negra degut al gran interès mundial en el seu cultiu i els avenços en tècniques moleculars, cosa que ha proporcionat avenços importants en el coneixement de la seva ecologia. Les contribucions més rellevants fan referència als condicionants del seu desenvolupament i a la seva reproducció.
Una tòfona madura conté milions d’espores tancades en ascs en grups de 1-6. Cada espora és portadora de només un dels dos tipus de encreuament (MAT1-1 i MAT1-2), però dins de cada asca hi ha espores dels dos tipus. Les micorizes (que formen l’òrgan simbiòtic amb l’arbre) també posseeixin només un dels dos tipus d’encreuament (mating types). La fertilització ocorre quan el miceli d’una micoriza (material matern) es troba amb un miceli del tipus oposat (material patern). No està demostrada la forma exacta de la contribució paterna; poden ser espores acabades de germinar de tòfones madures al camp, o dispersades per animals o insectes, o pot ser el miceli d’una micoriza del mateix arbre o d’un arbre veí.
Amb aquest descobriment, diversos estudis van analitzar la distribució dels dos tipus de creuament de les hifes i micorizes en plantacions de tòfones i en zones silvestres, productores i no productores de tòfones. Encara que s’han observat els dos tipus presents en un arbre, el patró més freqüent en camp mostra la tendència al fet que la majoria dels arbres propers dins d’una parcel·la presenten micorizes amb el mateix tipus de “mating type”. Aquesta observació destaca la importància de la introducció de material genètic: espores dispersades per animals i insectes o la gestió de la plantació amb aportacions d’espores o tècniques de maneig del sòl per posar en contacte els dos tipus de “mating types”.
García-Montero LG, Casermeiro MA, Hernando I, Hernando J. (2007a) Effect of active carbonate, exchangeable calcium, and stoniness of soil on Tuber melanosporum carpophore production. New Zealand Journal of Crop and Horticultural Science 35:139-146.
Linde C, Selmes, H. (2012). Genetic diversity and mating type distribution of Tuber melanosporum and their significance to truffle cultivation in artificially planted truffieres in Australia. Applied and environmental microbiology, 78(18), 6534–9. doi:10.1128/AEM.01558-12
Martin F, Kohler A, Murat C, Balestrini R, Coutinho PM, Jaillon O, Montanini B, Morin E, Noel B, Percudani R et al. (2010). Périgord black truffle genome uncovers evolutionary origins and mechanisms of symbiosis. Nature 464: 1033–1038.
Murat C, Rubini, A, Riccioni C, De la Varga H, Akroume E, Belfiori B, Guaragno, M, et al. (2013). Fine-scale spatial genetic structure of the black truffle (Tuber melanosporum) investigated with neutral microsatellites and functional mating type genes. The New phytologist, 199(1), 176–87. doi:10.1111/nph.12264
Paolocci F, Rubini A, Riccioni C, Arcioni S. (2006). Reevaluation of the life cycle of Tuber magnatum. Applied and Environmental Microbiology 72: 2390–2393.
Riccioni C, Belfiori B, Rubini A, Passeri V, Arcioni S, Paolocci F. (2008). Tuber melanosporum outcrosses: analysis of the genetic diversity within and among its natural populations under this new scenario. New Phytologist 180: 466–478.
Rubini A, Belfiori B, Riccioni C, Tisserant E, Arcioni S, Martin F, Paolocci F. (2011). Isolation and characterization of MAT genes in the symbiotic ascomycete Tuber melanosporum. The New phytologist, 189(3), 710–22. doi:10.1111/j.1469-8137.2010.03492.x
Rubini A, Belfiori B, Riccioni C, Arcioni S, Martin F, Paolocci F. (2011). Tuber melanosporum: mating type distribution in a natural plantation and dynamics of strains of different mating types on the roots of nursery-inoculated host plants. The New phytologist, 189(3), 723–35. doi:10.1111/j.1469-8137.2010.03493.x
Zampieri E, Rizzello R, Bonfante P, Mello A. (2012) The detection of mating type genes of Tuber melanosporum in productive and non productive soils. Appl Soil Ecol 57:9–15